1. 引言
神经科学告诉我们,大脑皮质各个局部空间的神经组织通过神经纤维,存在空间联系的规律性、渐变性、对应性。同时,大脑感觉中枢与身体各个局部空间的神经组织通过神经纤维,存在空间联系的规律性、渐变性、对应性 [1] 。这些联系特点表现在:神经纤维的联系是成束、规律的分布(如果不是规律性、渐变性的分布,神经纤维会纠缠在一起而不会是规律的成束);感觉刺激兴奋的感觉中枢的神经具有规律性、渐变性、对应性;人体“全息图”……。至于这种分布产生的原因,现在的学术界并没有相关理论解释。
根据胚胎发育的过程可知,神经纤维的这种对应性联系是神经元的突起的运动,特别是特殊突起(轴突)运动的结果。因而为了探讨这些理想规律,我们有必要讨论细胞和其突起的运动与分布情况。
对于机体的炎性细胞、肿瘤细胞的运动分布情况,由于与本文所讨论的胚胎环境条件的巨大差异,因而不在本文的讨论范围内,但空间结构基因的活性仍然会在它们的运动分布过程中发挥关键作用。
对于细胞的运动分布,学术界有许多相关研究 [2] [3] [4] ,但都相对初级,根本无法解释上述细胞的运动分布特点。为了更好地讨论细胞的运动分布特点,从而解释机体细胞,特别是神经细胞的这种对应性联系,我在本文引入了空间结构基因的概念。
空间结构基因在本文的讨论过程中起到了很好的中介作用,它将本文讨论的各个知识点有机地结合了起来。
关于空间结构基因与细胞的运动分布的关系,现在的学术界缺乏直接的研究。究其原因是我们对基因功能的研究远远不够,在现有的科技水平下,我们能够确定的是,空间结构基因的活性是细胞运动分布的关键因素,但无法明确具体有那些空间结构基因,它们在细胞运动过程中具体发挥了多大作用等等。因而我现在只能在“理想模型”下进行“理想”的“初级”讨论,并将讨论结果与客观事实相对比,从而尽可能保证讨论的相对准确性。
2. 空间结构基因
细胞的运动、静止是力作用的结果,由细胞产生的影响细胞自身运动的力主要有:使细胞突起产生的力、粘附的力、收缩的力。这些力有肌动蛋白、肌球蛋白相互作用产生的力,微管的延伸力,……等等,每种力(本文把由不同物质产生的力称为不同种的力,比如:肌动蛋白与肌球蛋白相互作用产生的力和由微管产生的延伸力之间便是不同的力)都是由一类相似的基本力组成的(何为基本力,我们知道产生一种力会有最小的物质要求,比如在某一正常环境下,如果肌动蛋白、肌球蛋白的量少于一个数目,肌动蛋白与肌球蛋白相互作用便不会产生影响细胞运动的力,我们把具有这一最少量的肌动蛋白与肌球蛋白相互作用而产生的力称为基本力),每类基本力可有多种(在不同的正常环境下肌动蛋白、肌球蛋白相互作用产生力所需要的最少量也不同)。细胞的运动与分布是由力引起的,力由基本力组成,基本力由相应的物质诱发(比如膜蛋白与细胞外物质的作用),这些物质又是由相应的基因或相应基因产生的物质产生的,我们把这些基因称为空间结构基因。
能够影响到细胞膜运动与黏附的因子都称为空间结构因子(比如肌动蛋白、某些信号分子等等 [6] )。因而,空间结构因子包含相关基因、dna结构,rna,蛋白质等等,而这些因子的核心是相关基因(比如与肌动蛋白对应的基因)的活性,这些相关基因就是空间结构基因。
细胞内物质的放射状运输、分布与空间结构基因
细胞内骨架系统对物资特别是高分子物资(体积相对较大)运动扩散的阻挡作用、微管对高分子物资的“放射状”运输、细胞内物资的代谢清除作用,使细胞内的某些高分子蛋白质及与蛋白质相关的物资具有从基因的转录、翻译、蛋白质的合成、合成后的运动分布具有一定的“放射状”对应性特点。这种细胞内物资的放射对应性运动分布应该是神经细胞维持其轴突结构功能相对稳定的重要因素(没有这种放射对应性运动分布,轴突应该无法保证一些在轴突分布的维持轴突结构功能稳定的高分子蛋白的浓度、活性的稳定)。这种放射对应性也使空间结构基因与细胞膜具有一定的放射对应性联系(前提是细胞核骨架系统及细胞质骨架系统的相对稳定),同理轴突与某些空间结构基因具有一定的放射对应性联系。比如移动细胞的β肌动蛋白的mrna,在其运动前端及后端的分布浓度的不同 [5] [6] ,显然无法完全用扩散来解释,而需要同时用这种放射状的运输来解释。又比如去除微管后,细胞无法维持极性 [6] 的实验现象,如果用细胞内骨架系统的相对稳定及微管的运输作用便能很好解释细胞的极性及细胞不同方向物资分布的差异了。
3. 细胞和其突起的运动、分布与空间结构基因活性
细胞与环境相互作用产生的力如果过小,则并不能影响细胞的空间位置,必须超过一个最小值才能对细胞的空间位置产生影响(这个最小值就是细胞运动的阻力,比如细胞与周围物资相互作用产生的某些阻碍细胞运动的粘附力)。同理,只有空间结构基因的活性变化达到一定程度,才能影响到细胞的空间位置,这种特点与皮质的柱状结构有什么关系?
3.1. 细胞的运动分布与空间结构基因
假设人类胚胎发育的早期,一类细胞的每个细胞的空间结构基因中,有a、b、c……这些空间结构基因,胚胎环境条件下,a基因产生的物质使这类细胞向南方运动,其他空间结构(b、c等等)基因使这类细胞向其他方向运动。在这类细胞的不同细胞中a空间结构基因的活性越低,其产生的运动力越小,当产生的运动力小于一定值时,细胞便不能运动,这个值便是a空间结构基因的最小运动力,那么便可能存在一群细胞,它们的a空间结构基因产生的运动力差别在最小运动力范围内,这群细胞分布在相同的一维空间内。假设仅a能够使这类细胞不同的细胞产生向南方的运动,运动的结果是这群细胞根据a基因的活性,顺南方逐渐分布。b、c等等基因的活性与细胞的分布关系也应该有类似a基因的这种渐变性。也就是说在一个环境中,在细胞数量足够多的情况下,充分运动的细胞具有与空间结构基因活性相关的空间渐变分布(胚胎时期的神经细胞就具有这样的特点)。
稍复杂一点的情况仍然存在这种渐变性:假设a1、b1、c1空间结构基因,它们都使细胞向南方运动,而它们在活性相同的情况下产生的力a2 > b2 > c2,假设有三群细胞在其它空间结构基因活性基本相同的情况下,a11细胞群的b1、c1空间结构基因没有活性,b11细胞群的a1、c1空间结构基因没有活性,……。这样,几个细胞群的可能分布是,a11细胞群在最南边,b11细胞群的南边部分与a11细胞群部分“重叠”,北边部分与c11细胞群部分“重叠”……。如。
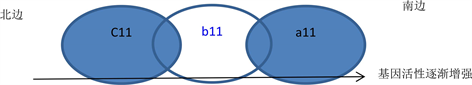
. the ideal model of cell spatial structure gene activity and cell distribution
. 细胞空间结构基因的活性与细胞的分布的理想模型
3.2. 神经细胞轴突的分布与空间结构基因
通过前面的讨论可知,理想环境、规则下,皮质局部神经元的分布具有与空间位置渐变相关的空间结构基因活性的渐变性。由于轴突对应的空间结构基因参与了胚胎时期神经细胞的运动,那么根据上面的讨论可知,轴突对应的空间结构基因的活性的渐变也会与轴突“起始”空间的空间位置的渐变相关、对应。
神经细胞的早期运动分布,主要是细胞之间的作用及一些分泌物质的影响 [4] 。神经细胞的轴突运动分布到某一局部,其运动分布是在力的作用下产生的,特点受到空间结构基因的影响,其运动分布特点仍然遵循上文关于细胞的运动分布的讨论,因而,运动分布到某一局部的一群轴突其对应的空间结构基因的活性的渐变也会与轴突“终止”空间的空间位置的渐变相关、对应。
理想状态下,轴突起始部位与这些轴突相关的局部空间的渐变性特点可以以某一函数集合表示,轴突对应的空间结构基因的活性的渐变性特点又可以用一函数集合表示,轴突终止部位与这些轴突相关的局部空间的渐变性特点可以以另一函数集合表示。我们可以得出,以轴突对应空间结构基因活性的渐变性特点为中介,轴突起始与终止部位的局部空间之间,存在规律的(函数集合)渐变相关性。这样,这两个空间通过神经纤维的联系,存在空间联系的规律性、渐变性、对应性特点。
简单来说,既然轴突在其起始空间与最终分布空间都具有空间位置渐变相关的空间结构基因活性的渐变性,那么以轴突的空间结构基因活性的渐变性为中介,这两个空间的纤维联系具有一定的规律性、渐变性、对应性也就是必然的了。
这些渐变相关性,只是理想模型下的一般讨论,实际情况复杂得多,但我们的神经系统也确实有这种空间位置联系的渐变性,也正好反应了神经细胞空间结构基因活性的渐变性与空间位置的渐变性相关。
4. 相似细胞空间结构基因活性的空间相似分布性
我们说两细胞的空间结构基因活性相似,是指这两细胞的空间结构基因的活性、在胞核内的空间分布、对应终产物的分布的相似。可以说具有相似起始点、且空间结构基因活性相似的两细胞的运动轨迹相似(因为它们与环境物资的作用力相似,当然空间结构基因活性不相似的两细胞也有可能有相似的运动轨迹),也就是说它们具有相似的空间分布。既然具有相似空间结构基因活性的细胞由于力的作用在局部的分布具有相似性(具有空间结构基因活性的渐变相似性),那么,分布到机体组织局部具有相似空间结构基因活性的轴突,在力的作用下其分布也应具有相似空间结构基因活性的渐变性,这种渐变性必然导致神经系统不同局部轴突联系规律的点与点的对应性。
理论上,在相同的外部环境下,空间结构基因活性相似并不表示其所有的基因表达活性就一定相似(如果有基因的表达不会影响空间结构基因的活性,就可能存在这种情况),但细胞的外形一定相似(细胞的外形是各种力作用的结果)。不过一般情况下细胞基因的表达与空间结构基因的表达应具有一致性。因为空间结构基因所包括的基因非常广泛,而两个细胞的空间结构基因活性相似则表示两个细胞对应基因的空间结构、细胞内环境等的相似(因为基因的活性与基因的空间结构及相关活性物资的活性、浓度、分布特点等等密切相关),所以这种情况下,空间结构基因相似,让人很难想象其它基因的表达不相似(我们的机体、相似性状的细胞一般有相似的局部分布特点,而一个局部也会有多种性状的细胞分布)。
机体整体、局部的细胞、轴突对应空间结构基因活性的分布特点
理想标准模型下,机体不同局部的细胞之间必然存在大量的空间结构基因活性的明显差异,同时也存在大量空间结构基因的活性相似。理论上各个局部的空间结构基因活性的明显差异一定程度上决定了,细胞在哪个局部分布,而大量相似的空间结构基因活性又会使各个局部细胞的分布具有空间结构基因活性的相似分布性。
在哺乳动物从产生胚胎到发育成人的过程中,细胞的空间结构基因的活性始终影响到细胞的空间位置的分布,从而使胚胎细胞空间分布特点是:机体整体细胞空间分布与局部细胞空间分布,局部细胞空间分布与其他局部细胞空间分布具有相似空间结构基因活性的相似分布性,而且这些相似性分布具有渐变性、规律性。
同理,神经纤维(轴突)的分布也具有这种整体与局部的相似性分布。轴突的起始及终止运动的根本原因是空间结构基因的活性,因而,这种相似性分布所产生的后果是皮质的一个局部发出的分布到另一局部的神经纤维,其分布特点是:组成局部的各个亚局部之间及与整体之间存在部分相似的纤维起始与分布特点,而使这两个局部的每个亚局部之间都会存在相对充分且相似的纤维联系。这种分布特点是给大脑皮质任何两个达到一定程度的刺激,它们之间都会建立一定的记忆联系的基础,这种分布与兴奋特点是大脑皮质并行记忆与存储的基础。我在《拟人智能的实现中》这本书中就是模拟这种分布与兴奋特点进行编程,从而实现并行存储与计算的 [7] 。
5. 小结
空间结构基因对细胞、轴突运动分布的决定作用及与各种功能基因的关系是理解大脑功能、机体细胞分布及神经纤维联系的钥匙。空间结构基因也是我们理解皮质柱状结构的产生、并行存储兴奋、任意记忆之间能够建立记忆联系、机体的对称性、两个大脑半球……等等现象的钥匙。
但现今科学界对空间结构基因的结构与功能的研究非常有限。因而本文只能在理想模型下进行初级、大概的推论,更进一步的讨论期待更多与空间结构基因相关的实验数据。本文通过“理想标准模型”讨论得出,大脑、机体细胞分布及神经纤维联系在空间上具有空间结构基因活性相关的相似性、规律性、渐变性、对应性分布特点。
当然,以上讨论的都是理想状态。现实中因为具有某些空间结构基因活性的细胞不能产生,或者产生后又无法生存,因而许多情况下,器官局部同类型的细胞在空间结构基因活性上可能不存在严格的渐变性。一般情况下,应该是打乱了的渐变性(大脑感觉中枢与机体的对应性联系就不是严格的渐变性联系)。
本文的讨论并不完美,肯定会引起许多质疑,但我希望本文的讨论能够起到抛砖引玉作用,从而吸引更多的学者投入相关研究。