1. 引言
中医学的奠基之作《黄帝内经》中阐述的重要穴位一般是由众多干细胞巢依一定规律排列组合形成的群落,不同种类干细胞巢的有序分布构成中医经络系统。引进板块概念主要反映干细胞巢基本稳定不变的静态成分,作者提出经络系统的高级结构单位为板块——一个个类似于伞状分布的腧穴群落,板块
中多能干细胞
及其成体干细胞
的干细胞巢模型
是完全相同的 [1] [2] 。在文献 [1] 与 [2] 的基础上,本文论述了满足可重复性、可操作性要求的
、
和
多能干细胞巢模型,通过对比动植物在细胞组织分化、物种进化树和生殖繁衍等方面的异同,将文献 [1] 中类比无脊椎动物推测的干细胞分化链
修改补充为有更多客观事实支撑的
。
2. 多能干细胞巢模型
目前,大多数胚胎学家接受修改后的生物重演律:个体胚胎发育重演了一系列与祖先胚胎相对应的阶段,即重演它们祖先胚胎的某些特征,特别是种系特征性发育阶段,后者是指脊椎动物早期发育的共同阶段的特征 [3] 。脊椎动物多能干细胞
及其成体干细胞
的干细胞巢,彼此离散地分布在组织深部的特定位置上,成体干细胞
分化产生定向干细胞,后者产生的各级分化细胞参与组织器官的构建及其新陈代谢。腧穴(干细胞巢)是构成经络系统的基本结构单位,《黄帝内经》开宗明义地提出经络是决死生、处百病的“灵枢”,后者的含义就是灵验的、灵妙的本源和中枢系统。在个体胚胎发育过程中,干细胞巢的主要细胞组合模型和干细胞巢群落有序分布(经脉和络脉)的演化,要比其所处的组织器官结构形态的演化具有更加明显的保守性,即与祖先胚胎相对比相同相似度更高,唯有如此,个体胚胎发育才能进行快速的重演。
干细胞与巢环境中的主要细胞等存在相互作用,通过外遗传方式使其暂时性获得相应层次基因组表达上的全能性,表现为干细胞在其干细胞巢中才能长期具有自我更新和分化能力,虽然远远不及胚胎干细胞
和
具有的全能性,但是同样可以理解为其相对于自身而言获得了全能性 [2] 。脊椎动物的成体干细胞有7 × 4 × 2 × 8 × n种:7为7个层次,4为4大基本组织(结缔组织、肌肉组织、神经组织和上皮组织),2为左右两侧对称,8为8类细胞,n为从低等级到高等级金字塔式累积叠加的n个进化等级(
,鱼类的进化等级
)。在物种进化史上,腔肠动物出现了4大基本组织的分化;扁形动物建立了左右两侧对称的体制;原腔动物实现了7个层次的真正分离;环节动物出现了“八类细胞”功能(
)第一次以
对分方式分配给2类细胞(
和
);软体动物发生了“八类细胞”功能的第二次
对分,分化产生4类细胞(
与
和
与
);鱼类完成了“八类细胞”功能的第三次
对分,分化产生8类细胞,表现为骨与软骨、交感神经与副交感神经、髓系细胞和淋巴系细胞等彼此独立出现。因为动物细胞都能不同程度地表达“八类细胞”功能对应的管家基因组群,所以物种进化史上四大基本组织、左右对称和七个层次三者依序独立产生和八类细胞功能的三次对分是完全不同的,后者不能成为构建干细胞巢的第四要素 [2] 。
六度分隔(six degrees of separation)理论是社交网络研究中最著名的理论之一:你和任何一个陌生人之间所间隔的人不会超过六个,也就是说,最多通过五个中间人你就能够认识任何一个陌生人 [4] 。这里,动物的细胞组织划分为七个层次,决定了多能干细胞
及其成体干细胞
的干细胞巢模型
中6的存在,六度分隔理论不再仅仅是一个数学猜想,如同四色定理(大脑新脑皮功能区的四色地图)一样 [1] 。
《黄帝内经》提出的十二经脉气血子午流注规律,以及对任脉与督脉、阴跷脉与阳跷脉等循行分布的描述,表明
多能干细胞巢模型
中的6,是由另外六个层次中同一侧的、隶属相同基本组织的、八类细胞中同一细胞类型的定向干细胞迁移而来,也就是说,在一般情况下,经络系统中诸板块的伞盖内并不需要存在相对应的
成体干细胞巢。
可重复性和可操作性是生物重演律的基本要求,
多能干细胞巢模型
中6的组合是以上述最直接的方式满足生物重演律的基本要求,并且成体干细胞分化产生的三种定向干细胞
能够平等地参与同一干细胞巢的构建,也就是说这里可能存在的组合数j ≤ 36并没有特别的意义;
多能干细胞巢模型
中6并不是分别隶属六个层次的、任意种类的(陌生的)定向干细胞,此情形可能存在的组合数是天文数字,不能对应于种系特征性发育阶段,只能对应于个体性特征发育阶段。
多能干细胞巢是由七个片层状细胞团结构——与七个层次一一对应——严格依序叠加组合形成,
多能干细胞主要在特定的片层中分化产生相应层次的
多能干细胞,
多能干细胞巢组合模型
中的
与
多能干细胞诞生地的片层状细胞团结构
一般是基本相同的 [2] 。与前文同理,根据十二经脉气血子午流注规律等可以确定,
不但对应于七个层次中的同一层次,而且在8分类细胞中选择相同的细胞分类,例如手足三阳经脉和手足三阴经脉分别是肌肉组织、结缔组织中第六类细胞的腧穴群,同为8分类细胞中的第六类细胞。易见,
中,1是唯一可以确定的,即为另一侧的对称者;3分别隶属于另外的三大基本组织,3 (存在左右两侧之分)可能存在的组合数仅仅是23 = 8。
板块
中多能干细胞
及其成体干细胞
的干细胞巢模型
是完全相同的,表明正是干细胞巢主要成分的特异性决定了板块
中x的特定取值。生物重演律决定了
干细胞巢模型
中的定向干细胞只能都隶属于相同等级即第x等级,这是组合数最少的、最简洁的情形;多能干细胞
在其干细胞巢中只能分化产生第一等级、第二等级乃至第x等级成体干细胞
,干细胞巢限定了多能干细胞
分化产生的最高等级成体干细胞为第x等级;一种多能干细胞
在生命体中至少能产生n个板块——第一板块
、第二板块
至第n板块
[1] 。
脊椎动物起源于无脊椎动物,前者的
多能干细胞巢组合模型为
,后者如软体动物和节肢动物的
多能干细胞巢组合模型为
。脊椎动物
多能干细胞巢组合模型
可以视为主要是由两个
多能干细胞巢交融组合形成 [2] ,倘若其中一个
多能干细胞巢组合模型被确定,则另一个
多能干细胞巢组合模型就能被唯一地确定,反之亦然,两者之间不存在组合的多样性,这是生物重演律等的必然要求。
在经络系统的板块中,伞盖内的
多能干细胞巢至少被3 1 = 4个外来的
成体干细胞巢所环绕,这些处于相同等级的
成体干细胞与该板块多能干细胞
隶属于同一层次,并且在8分类细胞中是相同的细胞分类,其分化产生的定向干细胞
是构成该板块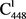 多能干细胞巢和
成体干细胞巢的主要细胞。
多能干细胞巢和
成体干细胞巢的主要细胞。
因为所处周围环境的区域性特征、自然选择的压力和生物重演律的要求等,脊椎动物
多能干细胞巢模型必须动态调整为
,相应的
多能干细胞巢模型必须动态调整为
,即七个层次中有选择性地挑选出y个层次
,其排列组合方式是固定不变的,前文所述的乃是无空缺的、最复杂的理想情形(
中的
)。因此,
多能干细胞巢至少被8y个外来的
成体干细胞巢所环绕,这些
成体干细胞都隶属于相同等级,并且在8分类细胞中是相同的细胞分类。
干细胞巢是一个复杂开放的微系统,作者认为,二种或二种以上定向干细胞竞争干细胞巢中同一角色(先增添、后删减)的自然选择过程,决定了个体性特征的形成与发展,在个体青少年时期特别明显;一种定向干细胞有可能兼具两个角色(1 1)或三个角色(1 1 1)的删减兼并过程,在个体中老年时期特别明显。
综上所述,脊椎动物的胚胎发育阶段,干细胞巢基本稳定不变的静态成分一般是高度有序的,上述
多能干细胞巢的
模型和
多能干细胞巢的
模型均是理想的标准模型;脊椎动物的胚后发育阶段,干细胞巢中诸外来的定向干细胞、成体干细胞乃至多能干细胞和胚胎干细胞等存在增添删减、更替兼并等形式的新陈代谢,大多数干细胞巢出现从井然有序到杂乱无序的熵增现象,导致干细胞巢中干细胞的更新分化能力逐渐衰退 [5] 。
3. 动植物进化树和细胞组织的分化
详见文献 [1] 的第二章节,约定动植物中具有全能性特征的细胞用
表示。将动植物细胞的减数分裂解释为“四大基本组织”功能的分离,第一次分裂实现“结缔组织与肌肉组织”和“神经组织与上皮组织”两者之间的对分
,第二次分裂实现了结缔组织和肌肉组织之间、神经组织和上皮组织之间的独立分离即
。减数分裂产生的子细胞有的能够重新获得全能性,但其遗传基因组与亲本相比已经发生了变异。
海绵动物开始出现细胞分化,没有四大基本组织的分化,没有神经组织。腔肠动物出现了神经细胞,有四大基本组织的分化即
。海绵动物出现能分泌骨针的成骨细胞等,表明其实现了四大基本组织”功能的第一次对分即
,具有“结缔组织与肌肉组织”功能的过渡性细胞可以特化成为成骨细胞等。在动植物进化树上,与海绵动物门存在对应关系的是真菌门中的鞭毛菌亚门和接合菌亚门(有性孢子2n),与腔肠动物门存在对应关系的是真菌门中的子囊菌亚门和担子菌亚门(有性孢子n)。
扁形动物出现了三胚层,获得了左右两侧对称的体制,干细胞分化链为
。原腔动物有口有肛门,具有完全的消化系统等,表明七个层次实现了真正地分离,es胚胎干细胞分化产生eg胚胎干细胞,第一条干细胞分化链为
。环节动物出现了“八类细胞”功能(
)第一次
对分产生2类细胞(
和
),第一条干细胞分化链为
。软体动物发生了“八类细胞”功能第二次
对分产生4类细胞(
与
和
与
),第一条干细胞分化链为
,其中
中包括左右对称的两种造血干细胞。节肢动物是无脊椎动物进化的顶峰,与软体动物相比较仅为过渡性细胞
的自然消失。
动植物进化树上,与扁形动物门存在对应关系的是绿藻门,植物的四大基本组织同样有
左右之分,因为植物细胞具有脱分化、再分化等内禀特征,所以左右之分只能在微观区域表达。与原腔动物门存在对应关系的是红藻门,注意植物细胞组织没有七个层次之分,这是植物细胞具有潜在全能性的原因。与环节动物门存在对应关系的是褐藻门,在此进化阶段,动物四大基本组织出现了“八类细胞”功能第一次
对分(
和
),植物四大基本组织“四类细胞”功能(对应于动物的
,约定用
表示)保持不变。与软体动物门、节肢动物门存在对应关系的是苔藓植物门,软体动物出现“八类细胞”功能第二次
对分,苔藓植物相应地出现“四类细胞”功能第一次
对分产生2类细胞(
与
)。
鱼类完成了“八类细胞”功能第三次
对分产生8类细胞,例如造血干细胞分化产生髓系祖细胞和淋巴系祖细胞,第一条干细胞分化链为
(
,鱼类的进化等级
),至此,构建动物组织器官的基本单位——体细胞——才获得稳定不变的结构功能形态。蕨类植物门中石松亚门的
,完成了“四类细胞”功能第二次
对分产生4类细胞,例如射线原始细胞分化产生韧皮射线细胞和木射线细胞,至此,植物组织器官的基本单位——体细胞——同样获得相对稳定不变的结构功能形态。
越是高等级的维管植物,越能清楚地观察到植物细胞组织的分化。植物的四大基本组织中,与动物上皮组织对应的是保护组织:四类细胞内的第八类细胞为表皮细胞等,第七类细胞为表皮中的保卫细胞、周皮补充组织的薄壁细胞和通气组织的薄壁细胞等,第六类细胞为韧皮薄壁细胞,第五类细胞为木薄壁细胞。与动物神经组织对应的是机械组织:第八类细胞为厚角组织细胞,第七类细胞为厚壁组织的石细胞,第六类细胞为厚壁组织的韧皮纤维,第五类细胞为厚壁组织的木纤维。与动物肌肉组织对应的是输导组织:第八类细胞为韧皮射线细胞,第七类细胞为木射线细胞,第六类细胞为筛管与筛胞,第五类细胞为导管与管胞。与动物结缔组织对应的是薄壁组织:第八类细胞为皮层细胞和同化组织的薄壁细胞等,第七类细胞为传递细胞,第六类细胞为贮藏组织的薄壁细胞、髓射线细胞等,第五类细胞为贮水组织的薄壁细胞等。注意,上述四大基本组织中产生分泌物的有关细胞或特化的细胞组合,总称为分泌结构,在细胞组织分化上没有基础性意义。
在动植物进化树上,两栖类动物的进化等级
,与其存在对应关系的是蕨类植物门中的真蕨亚门等;爬行类动物的进化等级,与其存在对应关系的是裸子植物门中的苏铁纲等;鸟类的进化等级
,与其存在对应关系的是裸子植物门中的买麻藤纲等;哺乳类中有袋类动物的进化等级
,与其存在对应关系的是被子植物门中的双子叶纲;哺乳类中真兽亚纲如灵长类等拥有全部的七个进化等级,与其存在对应关系的是被子植物门中的单子叶纲。
动植物细胞组织的分化及其dna遗传基因组的构成和演化等存在巨大的差异,从协同进化、平行进化等角度不能解释上述的一一对应关系,例如爬行类和鸟类归类为羊膜卵动物,这与裸子植物的分类命名完全相同。作者认为,遗传基因划分为硬件基因和软件基因两大类,硬件基因组集中分布在染色体dna等上,可以为有机体中全体细胞所共有,软件基因组离散地分布在细胞质和细胞核中,软件基因组的多少及其开启和关闭状态决定了动植物细胞种类和细胞分化 [1] ;动植物软件基因组的数量及其属性,有着基本相同的进化产生规律——以“斜坡”方式累积递增 [1] ,这是动植物进化树之间建立一一对应关系的物质基础。
4. 花的起源与进化
类比脊椎动物的第一条干细胞分化链,维管植物孢子体世代的细胞分化链表示为
,简略说明如下:
约定表示全能的胚细胞,类比动物的成体干细胞;
约定表示一一对应于叶(i)、茎(ii)和根(iii)的三种胚细胞,其区别在于表达最外层斜坡上处于进化阶段的软件基因组i、ii和iii时存在强度上的差异,
类比动物的定向干细胞,极易脱分化成为
;
表示
分化产生四大基本组织的细胞;
表示
进一步分化产生互为左右对称的子细胞;
表示
分化产生
和
两类细胞;
表示
分化产生八与七和六与五四类细胞,总共分化产生32种细胞;
和
对应于原分生组织,
、
和
对应于初生分生组织,
对应于次生分生组织。
蕨类植物为隐花植物,有营养叶和能育叶以及异型叶和同型叶等的划分,能育叶与营养叶之间的区别主要是边缘分生组织的有无,两者叶原基中的细胞分化链
是完全相同的。将植物细胞分化链中的
左右之分与动物第一条干细胞分化链中的
层次之分相类比,可以给出两条全新的细胞分化链:约定用w表示左右对称者实现
分离,用a表示“四类细胞”功能第一次
对分,用b表示“四类细胞”功能第二次
对分,这里只能给出三种排列组合顺序wab、awb和abw。作者认为,蕨类植物的能育叶为wab;裸子植物的能育叶为awb,其蕨类植物祖先的能育叶wab失去生殖能力;被子植物的花蕊为abw,花瓣为awb,花萼为wab。注意,在三条干细胞分化链的终端分化细胞
内,基因转录表达调控的方式是不同的,因为乘法交换律a × b = b × a不一定成立,植物细胞中产生不同的蛋白质复合体(例如由花器官发育基因的产物参与构成)等,因此必须重新认识控制花器官发育的abcde模型 [6] 。
能育叶中大孢子和小孢子的发生,显然必须对应于一条生殖细胞分化链,以被子植物为例,根据前文的论述,不难得到
和
以及
,简要说明如下:abw表示大孢子母细胞或小孢子母细胞及其前体细胞,
表示大孢子母细胞或小孢子母细胞经过减数分裂产生大孢子或小孢子
,后者恢复全能性转变为
,进入配子体世代,
分化产生对应于叶、茎和根的
,后者能够直接分化产生32种细胞中的任意一种,
在若干种细胞
的协同生长和营养保护之下,一个或者n个
全能态细胞转变成为
全能态细胞,后者发育成为生殖配子。
同理,作者认为裸子植物的生殖细胞分化链为
,配子体世代的细胞分化链为
和
;蕨类植物的生殖细胞分化链为
,配子体世代的细胞分化链为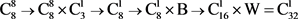 和
;苔藓植物生殖细胞分化链为
,苔藓植物配子体世代的细胞分化链为
和
,孢子体世代的细胞分化链为
;绿藻、红藻和褐藻等高等藻类孢子体世代的细胞分化链为
,生殖细胞分化链为
,绿藻、红藻和褐藻等高等藻类配子体世代的细胞分化链为
和
。
和
;苔藓植物生殖细胞分化链为
,苔藓植物配子体世代的细胞分化链为
和
,孢子体世代的细胞分化链为
;绿藻、红藻和褐藻等高等藻类孢子体世代的细胞分化链为
,生殖细胞分化链为
,绿藻、红藻和褐藻等高等藻类配子体世代的细胞分化链为
和
。
综上所述,植物孢子体与配子体的世代交替以及生殖器官(孢子叶和颈卵器)的发生和演化,从最基本的细胞组织分化角度来看,必须划分为被子植物、裸子植物、蕨类植物、苔藓植物和高等藻类(绿藻、红藻和褐藻)共五大类,高等藻类出现的同形世代交替和异形世代交替,维管植物孢子体世代(越来越复杂)和配子体世代(越来越简化)的进化趋势呈现出明显的此消彼长关系,特别是被子植物的花(花蕊)没有从其祖先裸子植物处寻找到渐进进化的线索 [7] ,得到了非常简洁的理论解释。
5. 动物经络系统中胚胎干细胞的分化
根据动植物进化树之间的对应关系,类比被子植物的生殖细胞分化链等,提出哺乳动物生殖腺内生殖细胞分化链为
,以及经络系统中第二条干细胞分化链为
。简要说明如下:
、
和
对应于“八类细胞”功能的三次对分,w表示左右对称者的分离,
对应于卵母细胞或精母细胞等,减数分裂产物
(与被子植物中的大、小孢子一样)首先恢复成为全能态细胞
,反之表明eg胚胎干细胞在经络系统中直接分化产生与减数分裂产物相对应的多能干细胞
,与第一条干细胞分化链
中的
完全相同。
同理得到,爬行类和鸟类的生殖细胞分化链为
,经络系统中的第二条干细胞分化链为
。鱼类和两栖类动物的生殖细胞分化链为
,经络系统中的第二条干细胞分化链为
。软体动物和节肢动物等的生殖细胞分化链为
,经络系统中的第二条干细胞分化链为
。
环节动物等的生殖细胞分化链为
,经络系统中的第二条干细胞分化链为
。原腔动物等的生殖细胞分化链为
,经络系统中的第二条干细胞分化链为
。扁形动物的生殖细胞分化链为
,经络系统中的第二条干细胞分化链为
。简要说明如下:将左右对称者分离和“八类细胞”功能的三次对分严格依先后顺序递减,从哺乳动物直至软体动物等,动物和植物遵循完全相同的细胞分化原理(实质是软件基因组的差异表达等);扁形动物的两条干细胞分化链
是完全相同的,说明了前述的递减操作应用于环节动物、原腔动物和扁形动物等是合理的;环节动物和原腔动物的生殖细胞分化链是相同的,这是因为es胚胎干细胞和eg胚胎干细胞都是全能细胞,也暗示了环节动物和原腔动物可能处于一个进化阶段的不同时期。
脊椎动物的经络系统中,es胚胎干细胞随机性存在于
、
干细胞巢之中,通过
诱导分化途径补充特定的
。eg胚胎干细胞随机性存在于所辖的
、
干细胞巢之中,通过
诱导分化途径补充特定的
[2] ,注意哺乳类动物如人类的经络系统中
。胚胎干细胞
和
的诱导分化以及迁移分布,使得经络系统成为一个开放进化的、自洽完备的生命信息系统。
构成干细胞巢的主要细胞为定向干细胞
和成体干细胞
,后者隶属于特定的进化等级,本文章节2中指出干细胞巢限定了多能干细胞
分化产生的最高等级成体干细胞为第x等级
,干细胞巢的进化等级特征决定了板块(经络系统的高级结构单位)是一个个彼此独立的、自洽完备的系统,板块
既不能自行退化降级为板块
,也不能自主升级进化为
,那么,以鱼类(
)进化为两栖类动物(
)为例,自然要问第3等级成体干细胞和板块
(及其对应的组织器官)是如何产生的?
经络系统中存在游离态的eg胚胎干细胞,不需要在任何干细胞巢中就能分化产生多能干细胞
,后者最初同样呈游离态存在于经络系统中,尤其是鱼类和两栖类动物的经络系统中并没有干细胞巢与之对应(
),游离态多能干细胞
分化产生的一部分多能干细胞
仍然能够保持游离态,后者分化产生n个进化等级的成体干细胞
,因此两栖类动物的第3等级成体干细胞得以大量产生,通过干细胞巢构成细胞的更新替换,依据叠加累积顺序将鱼类祖先中的一些板块
升级进化为两栖类动物的
板块。
众所周知,维管植物的原生分生组织分布在根与茎的顶端,叶片的生长与分化是叶原基的顶端分生组织与边缘分生组织和板状分生组织等交替活动的结果。在脊椎动物的经络系统中,胚胎干细胞
、
和
、
以及
多能干细胞巢一般富集不同等级的板块交汇处(类比植物的居间分生组织)以及板块的顶端(类比植物的顶端分生组织),针灸选取的重要腧穴集中分布在板块交汇处,例如在手足四肢的关节附近 [2] 。将由干细胞、干细胞巢等构成的中枢系统视为一种抽象的植物,将全能细胞
、
和多能干细胞
、
、
及其干细胞巢类比植物的分生组织,显而易见,经络系统的演变进化与维管植物的生长存在一定的相似性。
6. 讨论
脊椎动物的卵子或精子发育成熟而具有繁殖能力,可以类比于维管植物的大孢子或小孢子恢复全能性,这是否表示经络系统中
多能干细胞群必须能够分化产生全部的
多能干细胞?老年个体生殖腺的生殖功能自然衰退,是否表示经络系统中eg胚胎干细胞的更新分化功能自然衰退?是否暗示经络系统中es胚胎干细胞诱导分化产生多能干细胞
的功能同样自然衰退?《黄帝内经》曰:恬淡虚无,真气从之,精神内守,病安从来?
致谢
作者研究植物细胞组织分化、动植物进化树之间的联系以及被子植物花的起源演化时,与张建国有长期的交流讨论,在此表示感谢。